

Os principais fatores de estresse climático para as plantas são as altas temperaturas e a seca, que afetam o crescimento, o desenvolvimento e induzem mudanças morfológicas, fisiológicas e bioquímicas nas plantas (Fahad et al., 2017). Isso porque, cerca de 90% da massa das plantas é constituída por água, que atua em processos fisiológicos e bioquímicos essenciais (Taiz et al, 2017). As plantas demandam quantidade e distribuição adequadas de água ao longo do seu ciclo de desenvolvimento. A Figura 1 é um exemplo de uma lavoura de milho na safra 2021/2022, no município de São Vicente do Sul/RS. Essa safra teve influência do Fenômeno El Niño Oscilação Sul na fase La Niña, que no Rio Grande do Sul causa redução nas chuvas durante os meses de cultivo do milho (Noia Junior; Sentelhas, 2020).

Figura 1.Espiga de milho após déficit hídrico no município de São Vicente do Sul/RS, na safra 2021/2022.
Situações assim ocorrem quando o suprimento de água é inferior a demanda hídrica da planta, principalmente nas fases críticas. Em geral, os estágios iniciais do desenvolvimento das plantas são sensíveis ao déficit ou excesso de água no solo, afetando a produtividade das culturas, pois nesta fase está sendo definido um dos principais componentes de produtividade, o número de plantas por área. Déficit hídrico na floração e enchimento de grãos também leva a perdas significativas na maioria das culturas de grãos, isso porque nessas fases estão sendo definidos componentes de produtividade como número de grãos (dependente da polinização) e peso de grãos. Além de perdas na produtividade, o déficit hídrico pode reduzir a qualidade dos produtos agrícolas. Na Tabela 1, apresentamos a demanda hídrica durante o ciclo de desenvolvimento e os estágios críticos para o déficit hídrico em algumas culturas.
Tabela 1. Demanda hídrica durante o ciclo de desenvolvimento e estágio de maior demanda.
| Cultura | Demanda hídrica aproximada (mm/ciclo) | Época de maior demanda hídrica | Fonte |
| Café | 1200 | Floração e chumbinho | Carvalho et al. (2013); Gomes et al. (2007) |
| Soja | 450 a 875 | Floração e enchimento de grãos | Carvalho et al. (2013); Silva (2018); Tagliapietra et al. (2022) |
| Milho | 400 a 700 | Floração | Albuquerque; Resende (2007) |
| Citros | 600 a 1200 | Floração e formação do fruto | Magalhães et al. (2005) |
| Cana | 1000 a 2000 | Crescimento vegetativo | Carvalho et al. (2013); Abreu et al. (2013) |
Algumas estratégias podem ser adotadas para evitar o déficit hídrico, entre elas, o uso de irrigação suplementar, a escolha da época de semeadura das culturas e a escolha da cultivar ou híbrido. Por exemplo, Tagliapietra et al. (2022) identificou que a demanda por água durante todo o ciclo de desenvolvimento da soja no Sul do Brasil varia de acordo com o Grupo de Maturação Relativa (GMR), sendo necessários 730 mm, 850 mm e 875 mm, respectivamente para cultivares de GMR ≤ 5.5, GMR 5.6 a 6.4, GMR ≥ 6.4, completarem seu ciclo de desenvolvimento e produzirem 100 sc/ha. Ou seja, cultivares de ciclo mais curto necessitam menos água para atingir a mesma produtividade de cultivares com ciclo mais longo. Outro exemplo, é o sistema soja-milho no Centro-Oeste do Brasil, no qual, deve-se ajustar a semeadura da soja para o início do período das chuvas, utilizando cultivares de ciclo curto, para que seja possível o cultivo do milho dentro da janela recomendada pelo zoneamento agrícola, evitando o déficit hídrico (Pilecco et al., 2024).
No entanto, essas estratégias são definidas antes do início da safra. Após a semeadura da lavoura, a nutrição das plantas desempenha um papel fundamental na mitigação dos impactos do déficit hídrico. A disponibilidade adequada de nutrientes melhora a adaptação das plantas a condições adversas, reduzindo os efeitos negativos do estresse hídrico sobre a produtividade (Waraich et al., 2011). A nível global, a combinação de problemas relacionados a baixa fertilidade dos solos e estresses ambientais é a principal responsável por perdas na produção agrícola (Cakmak, 2005). Assim, cuidar da saúde do solo e da nutrição de plantas é fundamental para a estabilidade e produtividade das culturas, mitigando efeitos de estresses bióticos e abióticos, como o déficit hídrico.
A redução na produtividade causada pelo déficit hídrico pode ocorrer devido ao fechamento dos estômatos da planta, visando reduzir a perda de água através da transpiração (Taiz et al., 2017). Com o fechamento dos estômatos, a assimilação do CO2 atmosférico é reduzida, causando diminuição nas taxas fotossintéticas e, por consequência, na produção de biomassa. Além disso, em situações de elevado estresse, ocorre superprodução de espécies reativas de oxigênio (EROs). O acúmulo dessas espécies reativas danifica os componentes celulares, podendo levar à morte celular programada (Taiz et al., 2017). Para degradar as EROs a planta usa antioxidantes enzimáticos e não-enzimáticos, o que tem alto custo energético, comprometendo o crescimento vegetal e a produtividade. No entanto, com a redução da transpiração, a absorção de nutrientes do solo também é limitada. Por isso, garantir um suprimento adequado de nutrientes desde o início do ciclo é essencial. Diante desses desafios, vamos conhecer o papel dos principais macronutrientes, micronutrientes e aminoácidos na mitigação dos efeitos do déficit hídrico nas plantas.
O nitrogênio (N) é absorvido pelas plantas principalmente como nitrato e amônio, sendo o nitrato a forma mais disponível no solo. Dentro da planta, o nitrato é convertido em nitrito e incorporado em compostos orgânicos, processo mediado pela enzima nitrato-redutase (Taiz et al., 2017). Em condições de déficit hídrico, a atividade dessa enzima é reduzida (Azedo-Silva et al., 2004), mas a suplementação de N pode aumentar sua atividade e a absorção de nitrato. Além disso, a aplicação de N quando há boa disponibilidade de radiação solar fortalece os mecanismos antioxidantes da planta.
O fósforo (P) desempenha um papel fundamental no crescimento radicular e na regulação do potencial hídrico das folhas, auxiliando na manutenção da turgidez celular e na abertura dos estômatos (Taiz et al., 2017). Esse efeito contribui para a eficiência da fotossíntese, mesmo sob condições de déficit hídrico. O potássio (K) auxilia na redução da produção de espécies reativas de oxigênio (Hasanuzzaman et al., 2020) e melhora a fotossíntese em condições de estresse hídrico (Raza et al., 2014). Além disso, promove o desenvolvimento radicular, ampliando a superfície de absorção de água (Romheld; Kirkby, 2010).
O cálcio (Ca) tem um papel importante na recuperação de plantas submetidas ao estresse hídrico, contribuindo para a estabilidade das membranas celulares e atuando como mensageiro na sinalização de estresse (Taiz et al., 2017). Além disso, auxilia na ativação da ATPase da membrana plasmática, que regula o transporte de íons e contribui para a manutenção do balanço osmótico em células danificadas. O magnésio (Mg) estimula o crescimento radicular, aumentando a absorção de água e nutrientes (Hansel et al., 2021). Também melhora o transporte de açúcares das folhas para as raízes e reduz os danos causados pela oxidação luminosa nos cloroplastos, o que mantém a taxa fotossintética e a produtividade sob estresse (Waraich et al., 2011). Além disso, eleva a atividade de enzimas e moléculas antioxidantes, diminuindo os níveis de EROs (Waraich et al., 2011). O enxofre (S) contribui para a formação de cisteína, aminoácido importante para a desintoxicação de EROs (Hansel et al., 2021). Micronutrientes como ferro (Fe), cobre (Cu), manganês (Mn) e zinco (Zn) atuam como cofatores de enzimas antioxidantes, essenciais na eliminação de EROs (Jaleel et al., 2009).
Os aminoácidos são componentes essenciais ou participam da síntese de enzimas, proteínas estruturais, clorofila e hormônios vegetais, além de contribuírem para o transporte e o armazenamento de nutrientes nos tecidos vegetais. Desempenham, portanto, um papel fundamental no crescimento e desenvolvimento das plantas, bem como em sua adaptação a estresses bióticos e abióticos. A prolina e a glicina, destaques no aminograma do AZOGEL (Figura 2) e do GELAMIN (Figura 3), matrizes orgânicas dos produtos da ILSA, também exercem papel importante na mitigação dos efeitos do déficit hídrico, contribuindo para a resiliência das plantas em condições adversas.
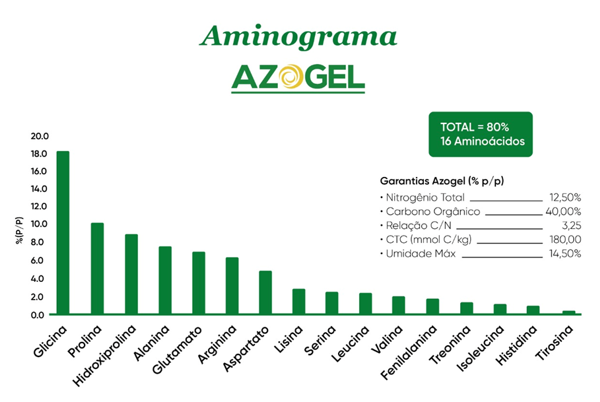
Figura 2. Aminograma do AZOGEL
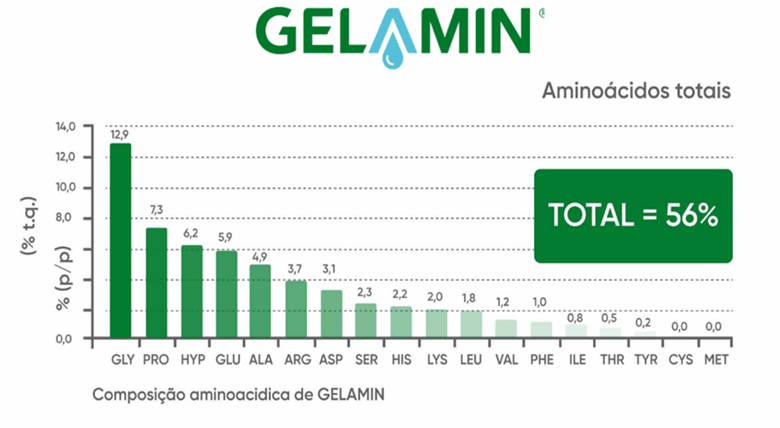
Figura 3. Aminograma do GELAMIN
Durante o estresse hídrico, a planta utiliza a prolina e a glicina betaína (composto derivado da glicina) para manter o equilíbrio osmótico, promovendo o acúmulo de solutos no citosol e no vacúolo das células. Dessa forma, a água (solvente) se move do meio externo (menor concentração de solutos) para o interior da célula (maior concentração de solutos), preservando sua hidratação (Taiz et al., 2017). A prolina e a glicina betaína são classificadas como solutos compatíveis, podendo ser acumulada em altas concentrações no citosol sem inibir a atividade de enzimas essenciais. Além da sua função osmótica, a prolina também atua como osmoprotetora, protegendo as plantas dos efeitos das EROs e servindo como fonte de carbono e nitrogênio durante a recuperação celular, quando as condições ambientais se normalizam (Taiz et al., 2017). A glicina, também atua na redução do estresse oxidativo, manutenção do metabolismo e na recuperação das plantas após o estresse hídrico.
Diante dos desafios impostos pelo estresse hídrico, a nutrição equilibrada das plantas é uma ferramenta que auxilia na redução dos efeitos negativos do déficit hídrico. Macronutrientes e micronutrientes desempenham papéis fisiológicos fundamentais nesse processo, como a regulação osmótica e a ativação enzimática, garantindo melhor adaptação das plantas em condições adversas. Tecnologias que combinam fontes minerais e orgânicas de nutrientes, além de serem fonte equilibrada de nutrientes e aminoácidos, possuem efeito estimulante ao desenvolvimento das raízes e benéficos ao solo, o que permite que as plantas explorem uma maior área de solo em busca de água e nutrientes. Assim, auxiliam na mitigação dos efeitos do déficit hídrico e possibilitam maior estabilidade para a produtividade das culturas. A ILSA oferece um portfólio completo para fortalecer as lavouras ao longo de todo o ciclo produtivo. Desde o tratamento de sementes, com ILSAMIN RADIX, até o suporte nutricional em diferentes fases do cultivo, com GRADUAL MIX (na forma sólida) e os fertilizantes líquidos ILSAMIN POTENTE e ILSAMIN FULL.
Referencias bibliográficas
ABREU, M.L. et al. Crescimento e produtividade de cana-de-açúcar em função da disponibilidade hídrica dos Tabuleiros Costeiros de Alagoas. Bragantia, Campinas, v.72, n.3, p.262-270, 2013.
ALBUQUERQUE, P. E. P. de; RESENDE, M. Irrigação: manejo de irrigação. In: CRUZ, J. C. (Ed.). Cultivo do milho. 3. ed. Sete Lagoas: Embrapa Milho e Sorgo, 2007.
AZEDO-SILVA, J. J.; OSORIO, F. F.; CORREIA, M. J. Effects of soil drying and subsequent re-watering on the activity of nitrate reductase in root and leaves of Helianthus annuus. Functional Plant Biology, v. 31, p. 611-621, 2004.
CAKMAK, I. Role of mineral nutrients in tolerance of crop plants to environmental stress factors. In: Proceedings from the International Symposium on Fertigation – Optimizing the utilization of water and nutrients. Horgen: International Potash lnstitute, 2005. p. 35-48.
CARVALHO, R.I. et al. Demanda hídrica das culturas de interesse agronômico. Enciclopédia biosfera, Centro Científico Conhecer – Goiânia, v.9, n.17; p. 2013.
FAHAD, S. et al. Crop production under drought and heat stress: Plant responses and management options. Frontiers in Plant Science, v. 8, p. 1147, 2017.
GOMES, N. M. et al. Crescimento vegetativo e produtividade do cafeeiro irrigado no sul do Estado de Minas Gerais. Revista Brasileira de Engenharia Agrícola e Ambiental, v.11, p.564-570, 2007.
HANSEL, F. D. Nutrição mineral como aliada das plantas na tolerância a estresses ambientais. Informações agronômicas, NPCT, n.9, 2021.
HASANUZZAMAN, M. et al. Reactive oxygen species and antioxidant defense in plants under abiotic stress: revisiting the crucial role of a universal defense regulator. Antioxidants, v. 9, p. 681, 2020.
JALEEL, C. A. et al. Antioxidant defense responses: physiological plasticity in higher plants under abiotic constraints. Acta Physiologiae Plantarum, v. 31, p. 427-436, 2009.
MAGALHÃES, A.F.J. et al. Sistema de produção para pequenos produtores de citros do Nordeste. Cruz das Almas: Embrapa Mandioca e Fruticultura Tropical, 2005. 178p. (Sistema de produção 17).
NOIA JUNIOR, R. S.; SENTELHAS, P. C. Yield gap of the double-crop system of main-season soybean with off-season maize in Brazil. Crop and Pasture Science, v.71, n. 5, p. 445-458, 2020.
PILECCO, I. B. et al. Ecofisiologia do milho visando altas produtividades. Santa Maria, Editora GR, 2024. 400p.
RAZA, M. A. S. et al. Exogenous application of glycinebetaine and potassium for improving water relations and grain yield of wheat under drought. Journal of Soil Science and Plant Nutrition, v. 14, p. 348-364, 2014.
ROMHELD, V.; KIRKBY, E. A. Research on potassium in agriculture: Needs and prospects. Plant and Soil, v. 335, p. 155-180, 2010.
SILVA, R. A. Impacto das mudanças climáticas sobre a produtividade e Pegada hídrica da soja cultivada na região do MATOPIBA. Tese de doutorado em Meteorologia, Centro de Tecnologia e Recursos Naturais, Campina Grande: UFCG, 2018.
TAGLIAPIETRA, E. L. et al. Ecofisiologia da soja visando altas produtividades. Santa Maria: Editora GR, 2022. 432 p.
TAIZ, L. et al. Fisiologia e desenvolvimento vegetal. 6.ed. Porto Alegre: Artmed, 2017. 888 p.
WARAICH, E. A. et al. Role of mineral nutrition in alleviation of drought stress in plants. Australian Journal of Crop Science, v. 5, p. 764-777, 2011.
Autores:
Eng. Agr. Msc. Isabela Bulegon Pilecco
Ing. Agr. Maestría en Ciencias. Thiago Stella de Freitas
Ing. Agr. Tuíra Barcellos



