
As raízes são estruturas essenciais às plantas e surgiram durante o processo de colonização do ambiente terrestre (OLIVEIRA, 2017; RAVEN e EICHHORN, 2014). Desempenham diversas funções, dentre elas o ancoramento e a sustentação, absorção de água e nutrientes, e a produção de hormônios reguladores de crescimento (VALVERDE‐BARRANTES et al., 2015). Além disso, as raízes são responsáveis por uma porção significativa da produtividade primária dos ecossistemas terrestres (MOMMER et al., 2015; MA e CHEN, 2016; FRESCHET et al., 2021), sendo importantes para o fornecimento de recursos para a comunidade microbiana do solo e influenciando, portanto, nos processos de decomposição microbiana e na ciclagem de nutrientes (FRESCHET e ROUMET, 2017; WANG et al., 2019). Por contribuírem com a melhoria dos aspectos físicos e químicos do solo, as raízes são consideradas como um bom indicador da qualidade dos solos (GARLET e SCHUMACHER, 2020).
O desenvolvimento radicular é um processo complexo, controlado por características inerentes à própria planta e ao ambiente no qual ela se encontra inserida (VALVERDE‐BARRANTES et al., 2015). Segundo Abramoff e Finzi (2015), um dos principais fatores que controlam a abundância e distribuição das raízes no solo é o genótipo das espécies que compõem a comunidade vegetal, entretanto as propriedades físicas e químicas do solo, tais como fertilidade, densidade, disponibilidade de oxigênio, pH, textura e temperatura, bem como a sazonalidade climática, também exercem forte influência (VALVERDE‐BARRANTES et al., 2015).
As raízes podem ser classificadas em finas e grossas (RIBEIRO et al., 2024). Os tecidos externos permeáveis e com baixo grau de suberização que compõe as raízes finas desempenham função essencial na absorção de água e nutrientes. Por outro lado, as raízes grossas atuam no suporte, condução de solutos, expansão do sistema de modo geral e fixação da planta no solo, sendo assim, crescem mais em profundidade (RATUCHNE et al., 2016). Autores como Correa et al. (2019) indicam que a qualidade do sistema radicular pode ser mensurada considerando a amplitude da superfície de contato do sistema com o solo, pois quanto mais profundo, maior o volume de solo explorado pela planta.
Relações entre crescimento radicular e condições físicas dos solos
As modificações provocadas pelo revolvimento do solo na estrutura, distribuição do tamanho dos poros e teor de carbono orgânico, alteram as forças de retenção de água no solo e sua disponibilidade, os quais são fatores determinantes para o desenvolvimento de plantas (SILVA et al., 2005). A conservação do solo e da água interfere na sustentabilidade do sistema produtivo, necessitando assim de várias práticas de conservação para preservação da qualidade física, química e biológica destes recursos naturais (MORAES, 2017).
Os macroporos são os principais poros responsáveis pelos fluxos de gases no solo (MORAES, 2017). A porosidade de aeração mínima de um solo deveria variar em função de diversos fatores, que determinam características de drenagem e o tempo que o sistema radicular fica exposto a uma oxigenação restrita (JONG VAN LIER, 2001).
Crescimento radicular e a interface solo-raiz
O aumento da extensão das raízes ocorre na região apical das raízes, e esta extensão dos eixos radiculares apicais bem como emissão de raízes laterais ocorrem para expansão da área de captação de água e nutrientes, além de potencialização da ancoragem das plantas (GREGORY, 2006). No ápice radicular são encontradas diferentes regiões da raiz (Figura 1), sendo que as divisões celulares ocorrem no meristema apical, a qual está confinada próximo da coifa que é um escudo de proteção das raízes, e pela zona de alongamento radicular (KONRAD, 2006), região onde ocorrem as pressões radiculares para penetração nos solos como resultado dos processos biofísicos da interação solo-raiz (BENGOUGH, 2006).
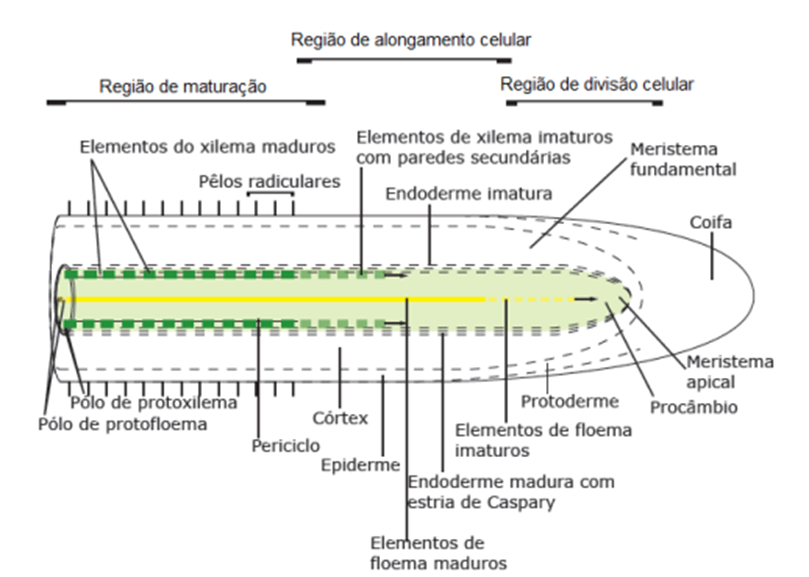
Figura 1. Representação esquemática do ápice radicular com regiões de divisão, alongamento e maturação celular. Fonte: Konrad (2006).
A divisão celular não resulta em aumento no comprimento radicular, mas sim fornece as matérias-primas para a expansão celular subsequente e, portanto, o crescimento radicular não é resultado somente da divisão celular (GREGORY, 2006). Na zona de alongamento, fora do meristema, as células aumentam de comprimento, acompanhadas por um grande aumento no tamanho do vacúolo e um aumento na área das paredes laterais da célula (GREGORY, 2006).
As limitações físicas dos solos afetam diretamente o crescimento radicular das culturas por meio da redução da taxa de alongamento radicular (BENGOUGH et al., 2011). Assim, caso não haja limitações químicas dos solos, as condições físicas dos solos durante o ciclo de desenvolvimento das culturas serão as responsáveis por permitir que as raízes possam se alongar e crescer no solo (MORAES, 2017).
O uso intenso de revolvimento do solo promove a dispersão das partículas de solo e causa o selamento superficial, promovendo entupimento dos poros do solo (VALENTINE et al., 2012), alterando os fluxos de ar e água no solo e do solo para as raízes das plantas. Assim, os limites físicos ao crescimento das plantas, são diferentes em solos com ou sem mobilização.
Influência da produtividade radicular no ciclo do Carbono
O ciclo global do carbono (C) é controlado basicamente pelo balanço entre a emissão e a absorção de CO2 pelos ecossistemas terrestres (PEREIRA JÚNIOR et al., 2016; MORANDI et al., 2021). A vegetação desempenha um papel fundamental na regulação desse ciclo, visto que a partir do processo fotossintético, uma parcela significativa de CO2 é absorvida e posteriormente estocada na biomassa vegetal (AGUIAR, 2018). Nesse sentido, sabe-se que de acordo com o ecossistema, as raízes podem ser responsáveis por até mais de 50% da produtividade primária (MA e CHEN, 2016; FRESCHET et al., 2021), superando a parte aérea em acúmulo de biomassa.
A matéria orgânica do solo pode representar grande parte do carbono total nos ecossistemas terrestres (NANZER et al., 2019). Assim, além de serem eficientes para a fixação carbônea, as raízes, principalmente as finas (por serem altamente dinâmicas), impactam diretamente o ciclo biogeoquímico do C (RATUCHNE et al., 2016).
Cabe salientar que a maioria dos estudos de biomassa nos ecossistemas terrestres se concentrou apenas no componente acima do solo, enquanto os reservatórios subterrâneos, como a biomassa radicular, ainda são bastante negligenciados (Pereira Júnior et al., 2016). A não incorporação desses estoques subterrâneos reduz a precisão das estimativas, impedindo uma melhor compreensão sobre a real contribuição dos diferentes ecossistemas terrestres no balanço global de carbono (RIBEIRO et al., 2024).
Influência das raízes finas na ciclagem de nutrientes
As raízes constituem uma importante fonte e sumidouro de nutrientes nos ecossistemas terrestres (RIBEIRO et al., 2024). Sua produtividade é fundamental para o forrageamento de nutrientes e absorção de água, ao mesmo tempo em que constitui uma entrada primária de nutrientes no solo por meio da sua renovação (LOIOLA et al., 2015). Segundo King et al. (2021), a influência das raízes na ciclagem de nutrientes pode variar de acordo com o seu diâmetro, sendo as raízes finas (≤2 mm) as que contêm significativamente maior concentração de nutrientes, tais como nitrogênio (N), fósforo (P) e magnésio (Mg), e consequentemente uma maior contribuição em relação as raízes mais grossas.
As raízes finas são responsáveis por uma grande parcela da produtividade primária dos ecossistemas terrestres, podendo em determinados ecossistemas superar a produtividade aérea (RIBEIRO et al., 2024). Diante disso, as concentrações de nutrientes presentes nas raízes finas podem ser maiores que as presentes nas folhagens das plantas e posteriormente na serapilheira (VERMA et al., 2021). Isso, associado ao fato de que as raízes finas são pouco longevas e apresentam uma acelerada dinâmica de produção, substituição e decomposição, fazendo com que elas constituam uma importante via de fluxo e ciclagem de C e nutrientes nos ecossistemas terrestres (VALVERDE‐BARRANTES et al., 2015; FRESCHET e ROUMET, 2017; GARLET e SCHUMACHER, 2020).
É importante enfatizar que o desenvolvimento das raízes no perfil do solo influencia significativamente a comunidade microbiana, principalmente por fornecer recursos orgânicos a partir da exsudação radicular (LANGE et al., 2015). Isso é de grande relevância, visto que, de acordo com a vegetação, a disponibilização desses recursos no solo pode favorecer o estabelecimento de relações simbióticas entre microrganismos e as plantas, o que possibilita um melhor contato e absorção de nutrientes essenciais, promovendo, consequentemente, um incremento na biomassa vegetal (FREIRE et al., 2020). Efetivamente os microrganismos do solo suprem o vegetal com nutrientes inorgânicos tais como N e P, e em troca, recebem nutrientes orgânicos oriundos da planta (WANG et al., 2019).
Visto esses fatores, fica evidenciado que a utilização de produtos ricos em Carbono e aminoácidos podem contribuir com a qualidade do solo e consequentemente melhora o enraizamento das plantas tornando a absorção de nutrientes mais eficiente.
A matriz Azogel presente em todos os granulados da Ilsa traz uma gama de opções que atendem a necessidade de vários cultivos e vários tipos e texturas de solo.
AZOGEL VS RIZOSFERA
A rizosfera (Figura 2) pode ser definida como a região do solo sob influência das raízes. O solo rizosférico tem características bem diferentes daquelas de um solo que se encontra distante das raízes, pois:
• Há grande presença de compostos orgânicos;
• Ocorrem inúmeros processos de simbioses específicas;
• O local possui baixa concentração iônica e baixo pH;
• Menor concentração de oxigênio.
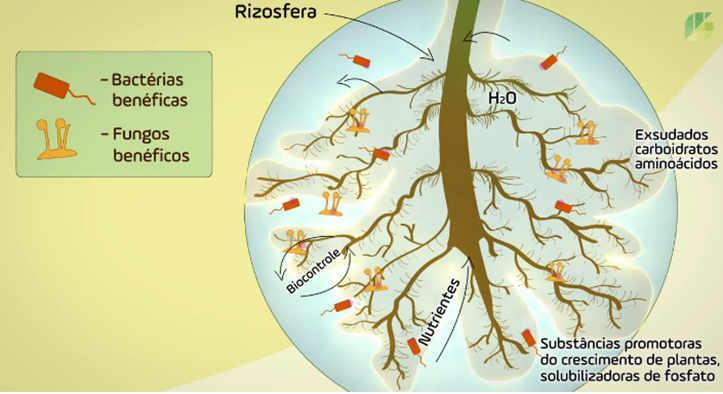
Figura 2. Mostra a área de atuação da rizosfera no solo. Fonte: De olho no material escolar.
De modo geral, o número de microrganismos presentes na rizosfera é muito maior do que em um solo não rizosférico e sua existência depende principalmente dos compostos orgânicos que a raiz exsuda para o solo. Por sua vez, a qualidade da rizosfera depende de uma alta disponibilidade de carbono orgânico. Desta forma, tanto o nitrogênio quanto os demais componentes orgânicos presentes em alta concentração no AZOGEL
contribuem positivamente para o desenvolvimento microbiano adequado na rizosfera. Este é um fator muito importante porque as bactérias se alimentam de carbono orgânico e as plantas se alimentam dos elementos que as bactérias e os fungos fornecem.
Como alternativa via TS (tratamento de semente), a ILSA conta com o que é um fertilizante exclusivo e de alta tecnologia da ILSA BRASIL, a base de GELAMIN®, que representa uma fonte natural de AMINOÁCIDOS de rápida absorção, que possuem ação tanto nutricional quanto estimulante dos processos fisiológicos das plantas. Sua moderna tecnologia de produção permite obter um produto único e de alta homogeneidade (sem variações de matérias-primas e tampouco nas garantias).
A aplicação de estimulantes via sementes é uma estratégia agronômica eficiente, focada em proporcionar um melhor início ao desenvolvimento das plantas. Esses produtos atuam diretamente no estímulo da germinação e na uniformidade da emergência, resultando em plântulas mais vigorosas.
O principal benefício é o crescimento melhorado do sistema radicular. Raízes mais profundas e ramificadas permitem às plantas explorarem maior volume de solo, otimizando a absorção de água e nutrientes essenciais para o seu desenvolvimento. Esse avanço no sistema radicular contribui para a formação de uma parte aérea mais robusta e saudávelAlém disso, raízes mais profundas aumentam a resiliência das plantas em condições adversas, como períodos de déficit hídrico. Essa característica é especialmente importante em regiões sujeitas à irregularidade de chuvas, promovendo maior estabilidade.
De maneira geral a ILSA, possui um portifólio completo para o bom desenvolvimento radicular das culturas, seja associando AZOGEL a suas formulações de base na linha GRADUAL MIX, ou de forma líquida associada ao tratamento de sementes.
Referencias:
ABRAMOFF, Rose Z.; FINZI, Adrien C. Are above‐and below‐ground phenology in sync. New Phytologist, v. 205, n. 3, p. 1054-1061, 2015.
AGUIAR, Diego Ribeiro de. Dinâmica e potencial de créditos de carbono na floresta manejada da Flona do Tapajós, Estado do Pará. 2018.
BENGOUGH, A. G. Root responses to soil physical conditions; growth dynamics from field to cell. Journal of Experimental Botany, Oxford, v. 57, n. 2, p. 437– 447, 2006.
BENGOUGH, A. G. et al. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. Journal of Experimental Botany, Oxford, v. 62, n. 1, p. 59–68, 2011.
CORREA, José et al. Soil compaction and the architectural plasticity of root systems. Revista de botánica experimental., v. 70, n. 21, p. 6019-6034, 2019.
FREIRE, Gabriel Araújo Paes et al. Dinâmica de serapilheira em uma área de floresta de terra firme, Amazônia Ocidental. Nativa, v. 8, n. 3, p. 323-328, 2020.
FRESCHET, Grégoire T.; ROUMET, Catherine. Sampling roots to capture plant and soil functions. Functional Ecology, v. 31, n. 8, p. 1506-1518, 2017.
FRESCHET, Gregoire T. et al. A starting guide to root ecology: strengthening ecological concepts and standardising root classification, sampling, processing and trait measurements. New Phytologist, v. 232, n. 3, p. 973-1122, 2021.
GARLET, Claudinei; SCHUMACHER, Mauro Valdir. Biomassa e comprimento de raízes finas em uma área de restauração florestal. Revista Brasileira de Gestão Ambiental e Sustentabilidade, v. 7, n. 15, p. 351-361, 2020.
GREGORY, P. J. Plant Roots Growth Activity and Interaction with Soils. Oxford, UK: Blackwell Publishing Ltd, 2006.
JONG VAN LIER, Q. DE. Oxigenação do sistema radicular: uma abordagem física. Revista Brasileira de Ciência do Solo, Viçosa, v. 25, n. 1, p. 233–238, 2001.
KING, William L. et al. The hierarchy of root branching order determines bacterial composition, microbial carrying capacity and microbial filtering. Communications biology, v. 4, n. 1, p. 483, 2021.
KONRAD, M. L. Transporte de nutrientes em plantas. In: BIZERRIL, M. X. A. (Coord.), (Ed.). Processos biológicos na captação e na transformação da matéria e energia: Módulo II. Brasília: Universidade de Brasília, 2006. p. 357– 383.
LANGE, Markus et al. Plant diversity increases soil microbial activity and soil carbon storage. Nature communications, v. 6, n. 1, p. 6707, 2015.
LOIOLA, Priscilla P.; SCHERER-LORENZEN, Michael; BATALHA, Marco Antônio. The role of environmental filters and functional traits in predicting the root biomass and productivity in savannas and tropical seasonal forests. Forest Ecology and Management, v. 342, p. 49-55, 2015.
MA, Zilong; CHEN, Han YH. Effects of species diversity on fine root productivity in diverse ecosystems: A global meta‐analysis. Global Ecology and Biogeography, v. 25, n. 11, p. 1387-1396, 2016.
MOMMER, Liesje et al. Diversity effects on root length production and loss in an experimental grassland community. Functional Ecology, v. 29, n. 12, p. 1560-1568, 2015.
MORAES, Moacir Tuzzin de. Modelagem do crescimento radicular de milho e soja sujeito a estresses hídrico e mecânico em latossolo. Universidade Federal do Rio Grande do Sul. Tese de Doutorado. 2017.
MORANDI, Paulo S. et al. Tree diversity and above-ground biomass in the South America Cerrado biome and their conservation implications. Biodiversity and Conservation, v. 29, p. 1519-1536, 2020.
NANZER, Marina Chiquito et al. Estoque de carbono orgânico total e fracionamento granulométrico da matéria orgânica em sistemas de uso do solo no Cerrado. Revista de Ciencias Agroveterinarias, v. 18, n. 1, p. 136-145, 2019.
PEREIRA, Lécio Resende et al. Carbon stocks in a tropical dry forest in Brazil. Revista Ciência Agronômica, v. 47, n. 1, p. 32-40, 2016.
OLIVEIRA, Paulo Marcelo Rayner. Controle do metabolismo e desenvolvimento da orquídea epífita Catasetum fimbriatum em resposta à incidência de luz no sistema radicular. 2017. Tese de Doutorado. Universidade de São Paulo.
RATUCHNE, Luis Carlos et al. Estado da arte na quantificação de biomassa em raízes de formações florestais. Floresta e Ambiente, v. 23, n. 3, p. 450-462, 2016.
RAVEN, P. H.; EVERT, R. F.; EICHHORN, S. E. Raven Biologia Vegetal. 8ª Edição. Rio de Janeiro, 2014.
RIBEIRO, Jéssica Araújo Heringer et al. Importância ecossistêmica das raízes: Uma revisão de literatura. Research, Society and Development, v. 13, n. 3, p. e0313345177, 2024.
SILVA, M. A. S. DA et al. Atributos físicos do solo relacionados ao armazenamento de água em um Argissolo Vermelho sob diferentes sistemas de preparo 1 Soil physics properties related to water storage and tillage systems in a Loamy Acrisol. Ciência Rural, Santa Maria, v. 35, n. 3, p. 544–552, 2005.
VALVERDE‐BARRANTES, Oscar J. et al. Aggregated and complementary: symmetric proliferation, overyielding, and mass effects explain fine‐root biomass in soil patches in a diverse temperate deciduous forest landscape. New Phytologist, v. 205, n. 2, p. 731-742, 2015.
VERMA, Abhishek K. et al. Fine root production and nutrient dynamics in relation to stand characteristics of chir pine mixed banj oak forests in central Himalaya. Flora, v. 279, p. 151808, 2021.
WANG, Jun-Jian et al. Long-term nitrogen addition suppresses microbial degradation, enhances soil carbon storage, and alters the molecular composition of soil organic matter. Biogeochemistry, v. 142, p. 299-313, 2019.
Autores:
Ing. Agr. Dra. Angélica Schmitz Heinzen
Ing. Agr. Maestría en Ciencias. Thiago Stella de Freitas
Ing. Agr. Tuíra Barcellos



